Impact of climate change effects on grapevine through a multi-year analysis in the Chianti Classico Area (Italy)
1 CREA—Council for Agricultural Research and Economics, Research Centre for Viticulture and Enology, Viale Santa Margherita, 80, 52100 Arezzo, Italy
2 Barone Ricasoli S.p.A. Società Agricola, Piazza Goldoni, 2 – 50123 Firenze, Italy
3 University of Cagliari, Department of Life and Environmental Sciences, University Campus, S.P. Monserrato-Sestu Km 0.700, Monserrato, 09042, Cagliari, Italy
*Corresponding author. E-mail: marco.ammoniaci@crea.gov.it
Abstract. Recent years have been increasingly characterized by the prevalence of extreme weather events due to climate change. Among these events, record-high temperatures and extended periods of drought are challenging the conventional viticulture techniques across many traditional grapevine-producing districts worldwide. The present study analyzes the data recorded over 16 years (2008-2023) in Chianti Classico, a renowned area in Tuscany (Italy) whose economy is based not only on the wine trade but also on the induced effects generated, such as wine tourism. The analysis correlated the historical climate patterns with the analytical profiles of the grapes at harvest. The results highlighted how increasing temperatures lead to an anticipation of the harvest date and, accordingly, a significant variation in grape chemical characteristics. This advance is linked mainly to achieving specific sugar concentrations in relation to the winery’s oenological objectives. As a result, organic acids and the phenolic fraction, along with their extractability, play a less decisive role and remain uncontrolled, potentially making the transformation process more challenging to manage.
Keywords: grape quality, time series analysis, ripening trends, Growing Degree Days (GDDs), Sangiovese.
The rise in temperatures on land, in the atmosphere, and in the oceans, combined with the retreating of snow and glaciers, which in turn contribute to rising sea levels, makes it hard to deny the phenomenon of global warming (IPCC, 2015). As these changes in the environmental balance challenge the sustainability of agriculture, valuable crops such as grapevines are not spared. Climate change, in fact, poses a risk to the cultivation of this plant in traditional growing areas and to the entire wine sector that depends on it (Al-Ghussain, 2019; Aydinalp and Cresser, 2008; Ortiz et al., 2021; Reidmiller et al., 2018). Vitis vinifera L., with 80 million tonnes produced annually over 7.3 million Hectares is the third most valuable agricultural product worldwide (OIV, 2020). Vitis vinifera (L.) is a remarkably resilient species, demonstrating impressive adaptability to extremely different climatic conditions (Duchêne, 2016). This plant, through human intervention (i.e., cultivar and clone selections, grafting, and agricultural practices), has been shaped over the centuries to thrive in different environments. The species of the genus Vitis that are of agricultural interest, thanks to their phenotypic plasticity, can be cultivated over a wide range of latitudes (50 °N to 40 °S), in 93 different countries and up to altitudes exceeding 3,000 meters (OIV, 2017; Van Leeuwen et al., 2024).
According to the current predicted possible climatic scenarios, the future is likely to hold a shift in terms of cultivation areas, with new regions becoming suitable for grape cultivation and others potentially at risk of being abandoned (Cameron et al., 2022; de Cortazar Atauri et al., 2017; Duchêne and Schneider, 2005; Van Leeuwen et al., 2019). As the novel climatic conditions move along the described pattern, grapevine phenology is altered, and while the length of the stages tends to remain constant, an anticipation of their occurrence is observed. Numerous studies have examined this phenomenon (Koch and Oehl, 2018; Dalla Marta et al., 2010; Tomasi et al., 2011), correlating climatic data with vine phenological stages across various production areas over the years, arriving at similar conclusions. Since the harvest date is primarily determined by the desired technological maturity of the grapes, climatic changes have resulted in an average advancement of 2-3 weeks in most wine-growing regions compared to past decades.
The current climate change in viticulture also has a strong impact on the vines’ physiological behavior during the vegetative season and, consequently, on the biochemical profile of the grapes (Van Leeuwen & Darriet, 2016). As a matter of fact, the vines spend part of the hot season under multiple abiotic stresses, with consequent impact on grape quality, as the latter is strictly linked to the environmental conditions (Drappier et al., 2019; Jackson and Lombard, 1993). An increased cluster exposure to solar radiation and a moderate water deficit can be desirable to boost the accumulation of polyphenols and aromatic compounds in the berries (Rienth et al., 2021; Van Leeuwen et al., 2022; Zarrouk et al., 2016). On the other hand, the adaptation of the plants to irregular hot and dry conditions might result in shrunk canopies and reduced photosynthetic activity, associated with withered clusters, possible sunburn damage and altered grape’s analytical profile (Cataldo et al., 2023; De Orduna, 2010; De Rességuier et al., 2023; Keller et al., 2016; Rustioni et al., 2023; Scholasch and Rienth, 2019). In particular, the worst effects of extended exposure of grapes to intense solar radiation and heat are lower acidic profiles, increased sugar accumulation (resulting in higher alcoholic content), and a general decoupling of phenological and technological maturity (Arrizabalaga et al., 2018; Kliewer, 1977; Mori et al., 2007).
Indeed, anthocyanins and sugar accumulation are closely linked during the early stages of ripening, but when the temperature gets over 35 °C, a significant slowdown in the biosynthesis of phenolic substances is often observed (Bergqvist et al., 2001; Gambetta and Kurtural, 2021; Pastore et al., 2017; Spayd et al., 2002). In most cases, when clusters are under intense heat, a delay in the onset of anthocyanin accumulation occurs, but once started, the rate of anthocyanin synthesis and accumulation remains constant (Sadras and Moran, 2012). Because of these unbalanced metabolic processes, when berries are long exposed to extreme temperatures, visible differences in berry size and color may be observed on the opposite sides of the clusters (Castellarin (a), et al., 2007; Castellarin (b), et al., 2007; Hernández-Montes et al., 2021). As the water stress reaches a specific threshold, the plant closes its leaf stomata to prevent damage from excessive water losses, consequently limiting the CO2 intake and the photosynthetic activity (Flexas et al., 1998; Medrano et al., 2003). As this occurs, sugar accumulation in berries is also impaired (Intrigliolo and Castel, 2010). However, under these conditions, berry growth is severely restricted as well, resulting musts display higher total final sugar concentration (Dai et al., 2009; Gambetta et al., 2020) due to a reduced berry weight. In this condition, to compensate for the energy deficit the plant speeds up the malate metabolism, causing a consumption of malic acid as fuel for the cells. As this happens, total acidity decreases, and pH rises in the musts at harvest (Hewitt et al., 2023; Sweetman et al., 2009; Van Leeuwen et al., 2009). Finally, water deficit may also advance the onset of sugar accumulationGiven the complex responses of Vitis vinifera at both plant and fruit levels, accurately assessing future possible scenarios for growing regions is crucial for planning mitigation strategies, as grape and wine production often represent the foundation of many agricultural communities. This study focuses on the shifts in grapevine ripening timing over the past 16 years in Chianti Classico, a key Italian wine district producing an average of 36 million bottles per year and valued at over 1 billion euros (Nesto and Di Savino, 2016). Focusing on Sangiovese, the Tuscan most emblematic variety, this research aims to assess the impact of rising temperatures on harvest timing and grape quality. By analyzing climatic trends, the findings will provide insights into preserving the distinctive characteristics of Sangiovese and suggest adaptive strategies for sustainable viticulture and enhanced wine production in this prestigious wine-growing area.
The study was carried out in the Chianti Classico district (Tuscany, Italy), in a 22 ha vineyard (43 ° 23 ‘ 02 “N, 11 ° 26 ‘ 04 “E, 320 meters a.s.l.) of Sangiovese (grafted on 110 Richter rootstock), over a 16-year period, from 2008 to 2023. The vineyard is located at the Barone Ricasoli Farm, the largest winery of the Chianti Classico district, having 240 ha of vineyards, displaying the pedological and climatological characteristics best suited for quality wine production. The farm has undergone several studies over the years, resulting in the creation of a zoning map. Moreover, the vineyard selected is the one closest to the weather control unit and has also been zoned, ensuring that all its soil characteristics are well-documented. The vines of the experimental vineyard are spaced 2.00 m between and 0.80 m within the rows, with parallel NW - SE orientation and an average 6 ° slope. The vines are trained on a vertical shoot positioned trellis, with spur cordon pruning (2 buds per spur, 4 spurs per vine). The soil has a clay-loam texture with high stoniness (up to 40 % limestone fragments) and a high calcium carbonate content (20-30 %). It has moderate water retention capacity, fast internal drainage due to rock fragments, low organic matter and nitrogen content, and a pH of 8.1-8.3. According to WRB classification, it is a Skeletic Calcaric Cambisol (Costantini, 2013).
The area has a predominantly temperate climate with hot-dry summer (Peel et al., 2007), characterized by wet winters and dry summers, with temperatures ranging from warm to extremely hot. During the growing seasons from 2008 to 2023, climate data were collected using an automated control unit (METOS ® by Pessl Instruments) located near the experimental vineyard. The parameters measured included daily maximum temperature (°C), daily average temperature (°C), daily minimum temperature (°C), and daily precipitation (mm), covering the period from April 1 to October 31, the conventional growing season for vines. Based on this data, the Growing Degree Days (GDDs) were calculated using the method described by (Winkler, 1974), with a temperature threshold of 10 °C. Essentially, GDDs represent the cumulative daily temperatures above a certain threshold that support active vine growth during the vegetative season. For each year, the focus was set on the growing period leading up to the Day Of Harvest (DOH, according to the Day-of-year calendar), assessing the GDDs from April 1 to the DOH, which varies in each single vintage (Table 1).
| Year | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | 2023 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DOH | 276 | 268 | 271 | 264 | 276 | 269 | 273 | 260 | 268 | 264 | 269 | 269 | 261 | 265 | 257 | 262 |
| 10/02 | 09/25 | 09/28 | 09/21 | 10/02 | 09/26 | 09/30 | 09/17 | 09/24 | 09/21 | 09/26 | 09/26 | 09/17 | 09/22 | 09/14 | 09/19 |
2.3. Grape maturity parameters
Every year, at DOH, 15 samples constituted of 200 berries each were randomly collected in the experimental vineyard and screened to assess the analytical parameters of the grape. Half of the berries were manually pressed, and the obtained juice was analyzed to assess the technological parameters: reducing sugars, pH, and total acidity. Analyses were directly performed on the filtered must samples using Fourier Transform Near-Infrared Spectrometry (FT-NIR) with the WineScan FT 120 system (Foss Italia S.r.l., Italy). All the above-mentioned parameters need to fall into specific ranges dictated by the Chianti Classico D.O.C.G. production regulations, in order to the wine to present the typical sensorial profile and claim the appellation. The second half of the samples were analyzed as described by Ribéreau-Gayon et al. (1965) and Saint Cricq de Gaulejac et al. (1998) to determine phenolic maturity indices: Total Anthocyanins, Extractable Anthocyanins, and the Cellular Maturity Index. All these indices are related to the phenolic composition of the wines allow to evaluate both their quantity and color. In detail, Total Anthocyanins are obtained by analyzing an extract prepared with a highly acidic buffer (pH 1.00) and represent the total anthocyanin content in the berries, expressed in mg/L of must. Extractable Anthocyanins, also expressed in mg/L of must, refer to the amount of anthocyanins measured in an extract obtained with a buffer at pH 3.20, similar to that of the wine. This fraction represents the bulk of pigments that are subject to the technological winemaking process. The Cellular Maturity Index reflects the ability of the berry skin to release anthocyanins and was determined using the following ratio:
Given the structure of the formula, lower index values correspond to a higher proportion of extractable anthocyanins, indicating a more advanced maturity stage.
The data collected and organized were analysed using the R program (R Core Team, 2023, https://www.R-project.org/) inside RStudio environmental. To evaluate the interactions between both weather and oenological parameters, a statistical analysis was performed using Pearson correlation coefficient (r) and evaluated its p-value. These two values together allow to understand how strong the relationship between two variables is, and whether it is statistically significant. These statistical results are shown within the graphs developed through the graphical tool ggplot2 (Wickham and Wickham, 2016).
The data processed from the weather station (Table 2) between April 1 and October 31 in 2008-2023 shown an average GDDs accumulation of 1907. The year with the lowest GDDs was 2010, with a total of 1605, while the highest was recorded in 2022, with 2136 GDDs. Over the same period, rainfall averaged 414 mm, with the lowest amount of water fed occurring in 2011 at 213 mm and the highest in 2016 at 586 mm. Events of temperatures dropping below 0 °C after 1st April (after bud burst) occurred in the following years: 2012 (2 days), 2015 (2 days), 2017 (2 days), 2020 (3 days), 2021 (4 days), 2022 (5 days), and 2023 (3 days), with all occurrences happening in the first half of April. The increasing frequency of late frosts after 1st April could increased the risk of damage to the newly sprouted vegetation. Additionally, GDDs data processed specifically from April to harvest day, revealed an average GDDs accumulation of 1692. Lowest annual GDDs over the years occurred in 2010 (1501 GDDs), and the highest was in 2017 (1842 GDDs). Mean precipitation for this period was 301 mm, with the lowest value in 2020 and the highest in 2010 with 135 mm 450 mm respectively. An additional metric of interest for long-term grape analysis was the frequency of days with temperatures exceeding 35 °C, which averaged 17 days per year.
| Year | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | 2023 | Average |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GDDs from 1 April to 31 October | 1763 | 1883 | 1605 | 1880 | 1917 | 1749 | 1748 | 1959 | 1877 | 2064 | 2076 | 1975 | 1913 | 1904 | 2136 | 2070 | 1907 |
| Rainfall (mm) from 1 April to 31 October | 476 | 411 | 523 | 213 | 474 | 539 | 399 | 429 | 586 | 421 | 417 | 325 | 261 | 219 | 477 | 460 | 414 |
| GDDs from April to DOH | 1600 | 1715 | 1501 | 1663 | 1758 | 1545 | 1555 | 1731 | 1685 | 1842 | 1831 | 1746 | 1714 | 1694 | 1831 | 1666 | 1692 |
| Rainfall (mm) from April to DOH | 393 | 316 | 450 | 157 | 308 | 304 | 382 | 273 | 435 | 408 | 345 | 247 | 135 | 181 | 199 | 282 | 301 |
| Yearly number of days Tmax > 35 °C | 9 | 13 | 1 | 12 | 23 | 6 | 0 | 25 | 3 | 40 | 7 | 21 | 19 | 20 | 37 | 31 | 17 |
| Yearly number of days Tmin < 0 °C | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 5 | 0 | 2 | 0 | 0 | 3 | 4 | 5 | 3 | 1.5 |
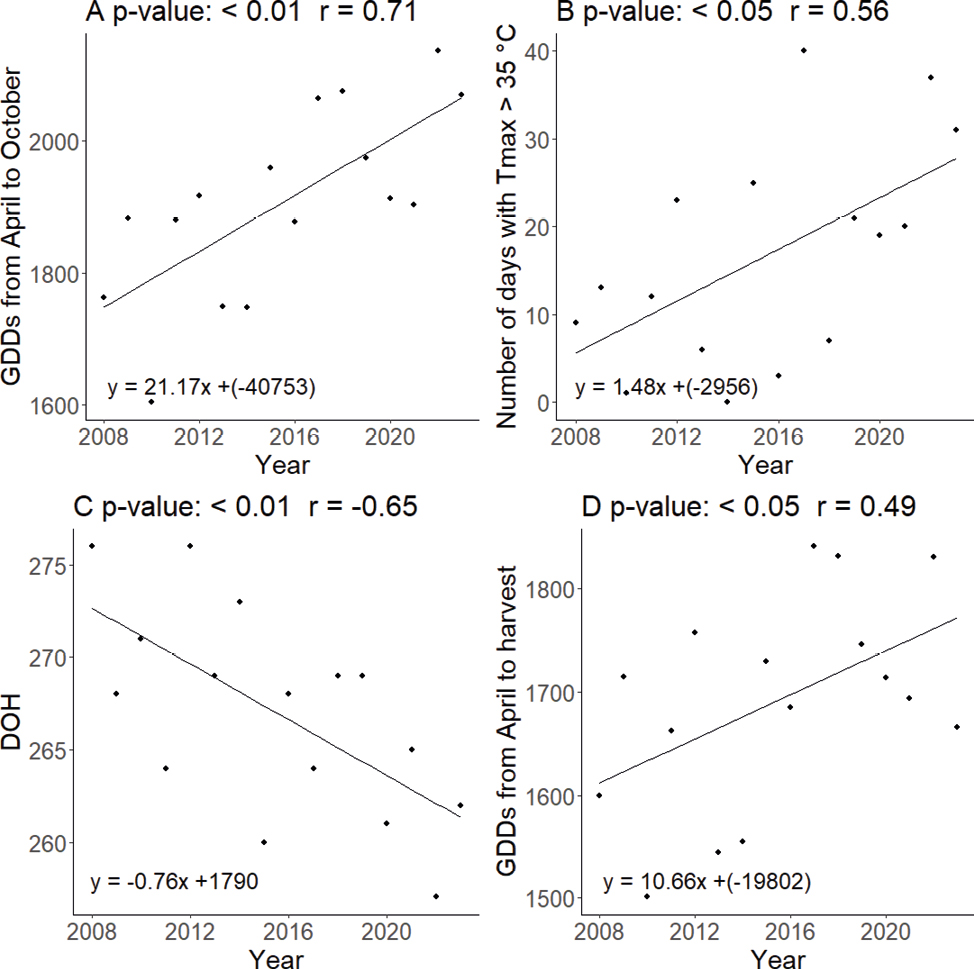
As shown in Figure 1, the correlation between the year and the GDDs from April to October (r =0.71, p-value < 0.01), suggested that, over the years, there had been a significant increase in the heat units accumulated during the growing season. With the rise in temperatures over the years, a direct correlation was observed between the year and the number of days with temperatures exceeding 35 °C (r =0.56, p-value < 0.05). The significant correlation between the year and the DOH was explained by the fact that as the years progressed, the harvest dates occured in advance (r =-0.65, p-value < 0.01), indicating a significant tendency. Despite the progressively earlier harvest, which reduced the number of days from April 1 to harvest, GDDs at DOH continued to increase (r = 0.49, p-value > 0.05).
As shown in Table 3, the oenological parameters obtained from grape samples collected at the DOH presented the following average values: reducing sugars, 234 g/L; pH, 3.27; and total acidity, 6.57 g/L. The year 2011 exhibited notably high levels of reducing sugars and low pH values, while 2016 and 2020 showed high levels of both sugars and pH. In contrast, the 2014 season recorded the lowest mean sugar concentration (209 g/L) and pH (3.13), along with the highest total acidity value (8.30 g/L). On the contrary, the lowest total acidity (5.20 g/L) was observed in 2020. Total Anthocyanin levels averaged 1093 mg/L, while Extractable Anthocyanins averaged 644 mg/L, resulting in a mean extractability of 41 %. Significant variations in anthocyanin levels were observed, with the higher concentrations noted in 2011, 2016, and 2020, while the lower levels were recorded in 2012, 2014, and 2017.
| Year | Reducing sugars (g/L) | pH | Total acidity (g/L tartaric acid) | Total Anthocyanins (mg/L) | Extractable Anthocyanins (mg/L) | Cell Maturity Index (%) |
|---|---|---|---|---|---|---|
| 2008 | 220 | 3.20 | 7.20 | 1162 | 516 | 56 |
| 2009 | 219 | 3.19 | 6.30 | 1199 | 613 | 49 |
| 2010 | 235 | 3.20 | 6.94 | 1059 | 643 | 45 |
| 2011 | 252 | 3.21 | 6.60 | 1289 | 891 | 31 |
| 2012 | 223 | 3.28 | 5.98 | 736 | 426 | 42 |
| 2013 | 228 | 3.22 | 6.87 | 1129 | 585 | 48 |
| 2014 | 210 | 3.13 | 8.30 | 817 | 478 | 41 |
| 2015 | 230 | 3.26 | 7.08 | 1002 | 659 | 34 |
| 2016 | 252 | 3.42 | 5.74 | 1583 | 873 | 45 |
| 2017 | 251 | 3.23 | 6.59 | 754 | 593 | 21 |
| 2018 | 243 | 3.37 | 6.49 | 1123 | 698 | 38 |
| 2019 | 238 | 3.30 | 6.37 | 933 | 565 | 39 |
| 2020 | 261 | 3.41 | 6.41 | 1359 | 842 | 38 |
| 2021 | 232 | 3.21 | 6.21 | 1066 | 557 | 48 |
| 2022 | 234 | 3.46 | 5.20 | 1140 | 678 | 40 |
| 2023 | 225 | 3.34 | 6.92 | 1112 | 689 | 38 |
| Average | 235 | 3.27 | 6.57 | 1093 | 644 | 41 |
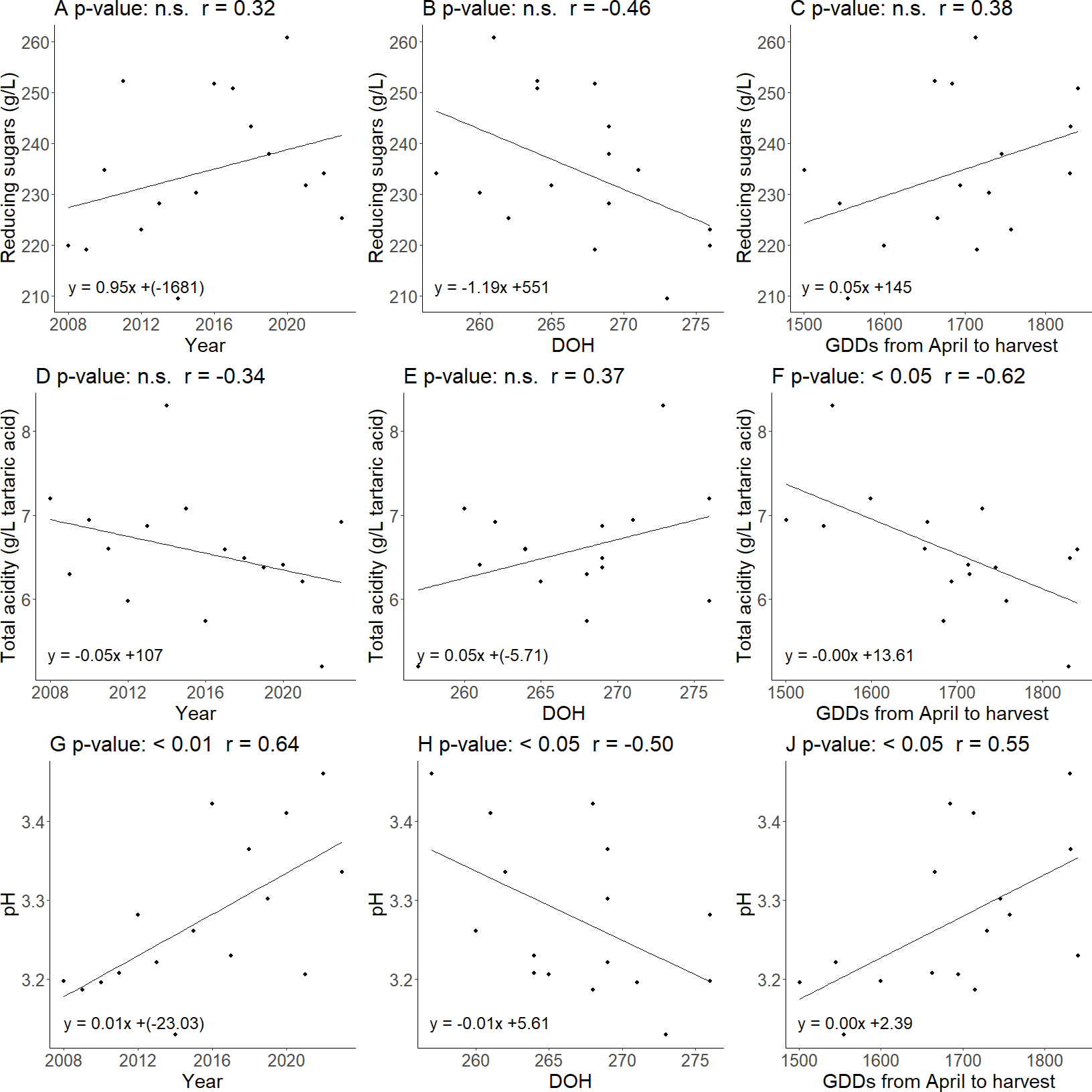
In Figure 2, although the correlation between reducing sugars and DOH was not statistically significant (p value = 0.07), it emerged that reducing sugars decreased as DOH increased, with a near-significant trend. The reducing sugars shown no statistically significant correlations with the other variables considered. A significant negative correlation was found between total acidity and GDDs (r = -0.62, p-value <0.05). For pH, both a positive trend over the years (r = 0.64, p-value <0.01) and with the GDDs (r = 0.55, p-value <0.05) are observed, while a significant negative correlation was found with DOH (r = -0.50, p-value <0.05).
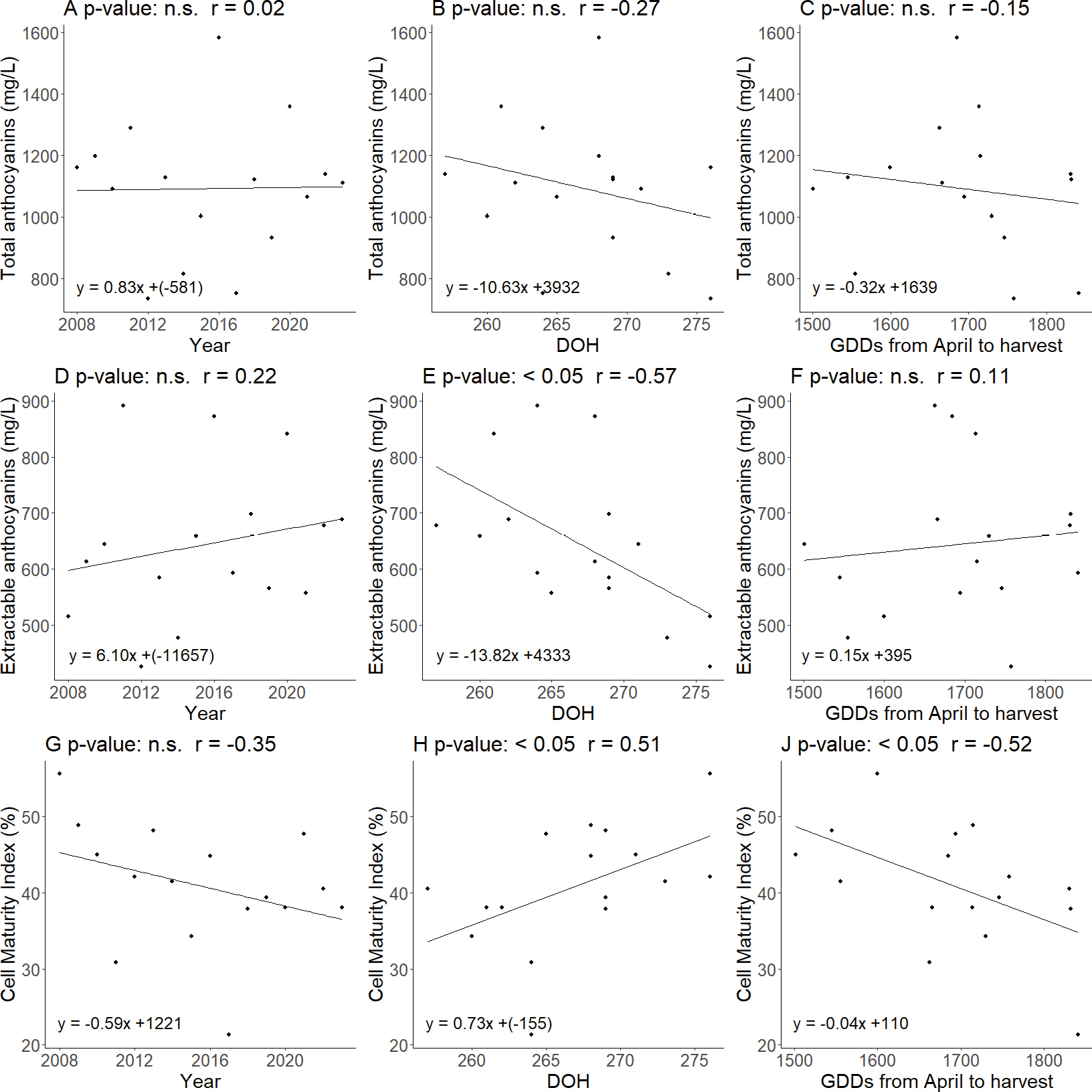
As illustrated in Figure 3, there was no correlation among Total Anthocyanins and the other parameters considered. Anyway, the extractable anthocyanins presented a significant correlation only with the DOH (r = -0.57, p-value <0.05), showing a decrease as this variable increased and indicating that a later harvest reduced the amount of Extractable Anthocyanins. Despite these results, a significant correlation was found between the Cell Maturity Index and DOH (r = 0.51, p-value <0.05), showing an increase with later harvest dates and decreasing GDDs at harvest (r = -0.52, p-value <0.05). This indicated that a late harvest promoted higher cellular maturation, while greater heat accumulation could reduce it.
The increasing frequency of enduring high temperatures and heat waves determines a growing economic impact on cultivated species, including Vitis vinifera.. In the area of investigation, Chianti Classico, a leading wine-growing district in the Italian panorama, the impact on grape quality (reducing sugar, acidity, pH, anthocyanins) in Sangiovese (the main black-berried cultivar in Italy) was evaluated over 16 consecutive growing seasons, from 2008 to 2023. The study here proposed helps to define what the possible future scenarios might be. Results highlight a significant rise in temperatures during the considered growing seasons (GDDs from April 1st to October 31th), with an average of 1649 GDDs from 2008 to 2012, leading to a 8 days advance in the harvest date compared to the period from 2019 to 2023, which had an average of 1729 GDDs. Along with the correlation between years and the increase in GDDs, there is an increase in days with extreme temperatures (>35 °C). To counter this trend, harvest is increasingly anticipated, which helps maintain relatively constant levels of total soluble solids and total phenolics at harvest. On the other hand, late frosts became more frequent after the 1st April , hence, late pruning was carried out to delay the vegetative awakening of the vines, without significantly compromising yield or grape quality (Poni et al., 2022).
Despite, GDDs show a tendency to continue to increase at DOH. This finds justification on the fact that temperature has a controversial effect on plant and cluster’s metabolism, where sugar and organic acids are both subject to anabolic and catabolic processes accordingly to the temperature’s range. Sugar accumulation is indeed most efficient in the range of 22 - 28 °C but comes to a complete stop when 35 °C is reached (Keller, 2020). Grapes directly exposed to extreme heat and solar radiation may experience a blockage in sugar accumulation, although berries in the shade remain active (Kliewer and Lider, 1968). In general, extreme heat interrupts ripening, slows physiological processes and extends the time required to reach maturity. Rising GDDs at DOH, combined with heat-induced interruptions, complicate ripening management by increasing thermal accumulation during the growing period, disrupting the balance between metabolic and ripening processes. Reducing sugars remain the main decision parameter to start the harvest, as supported by the relatively constant levels over time. This decision, however, often comes at the expense of other analytical parameters of the grapes.
For the above-described earlier occurrence of the date of harvest, a reduction of the total acidity measured on must is observed, when compared with GDDs at DOH. Also measured a higher pH as sun-exposed berries tend to have a significantly lower amount of malate (Kliewer and Lider, 1968; Reynolds et al., 1986). This phenomenon is likely to be caused by the reduced photochemical activity of the plants under heat stress, and the consequent use of malic acid as a source of energy by the plant (Ribéreau-Gayon et al., 2021). These increasing pH levels in the berries are likely to rise further, with potential impacts on the typicity of the final wine, as the number of extreme-heat-days continue to grow.
Concerning total anthocyanins in the musts, final concentrations have shown to remain relatively stable. Is indeed acquired that intense heat may decouple the onset of phenolics and sugar accumulation but, once started, phenolics synthesis might catch up. The lack of association between the total anthocyanin contents and the other parameters detected can be therefore explained by the fact that total phenolic substances are influenced more by genetics rather than by climate except in the case of prohibitive environmental conditions. This fact is supported by other authors who have correlated the phenological stages to a constant amount of GDDs in different grape cultivars (Zapata et al., 2015). In this respect, it was significant to observe that the general trend of grapes harvested earlier produced musts with higher amounts of extractable phenolic compounds due to the possible effects of the intense heat and light exposure on berry tissues. Anthocyanin production peaks at an optimal berry temperature of approximately 30 °C but declines when temperatures exceed 35 °C (Kliewer, 1977; Spayd et al., 2002). Prolonged exposure to excessive heat and excessive light radiation can induce oxidative stress, which not only inhibits anthocyanin synthesis but also contributes to its degradation (Mori et al., 2007). The Cell Maturity Index confirms that the percentage of extractable anthocyanins is higher in the grapes harvested earlier. Moreover, the significant correlation between GDDs to harvest highlights the influence of climate on grape maturity. This supports the observation that warmer conditions during the vegetative growth phase led to higher extractable anthocyanin percentages.
The data collected over a prolonged period, 16 years (2008 – 2023), analyzed in this study allow us to conclude that there is evidence of an overall increase in temperatures during the grapevine growing season, leading, in general, to an earlier harvest date within every year. Events of extreme heat are also occurring with increasing frequency. Sugar level remains the main factor in determining the harvest day for a winery, while total acidity has been decreasing over the years. While the grape total anthocyanin content has not significantly changed over time, the concentrations of extractable anthocyanins increased, likely due to the grapes experiencing more thermal stress.
Advance the harvest impacts the balance of sugars, acidity, and aromas, altering the wines’ flavour profile, and at the same time, it brings on logistical and economic challenges, such as managing seasonal labour and adapting production cycles.
Although the advancement of the harvest in response to the accelerated plant’s metabolism is mainly driven by the need to maintain constant the sugar level, to predict a future scenario remains difficult as extreme-heat-days are also occurring more frequently. As plant physiological processes are an articulated system, and different metabolic pathways get involved in response to combined stresses, is rightful to assume, as one of the possible scenarios, that extreme heat events may prevent further advancements delaying the ripening by halting photochemical activity. This underscores the complex relationship between climate change and agricultural practices, emphasizing the need for sustainable farming methods (e.g., optimized irrigation, canopy management, shading nets, reflective clays) and ongoing research to address these challenges while preserving the wine sector’s quality and stability.
The authors want to thank the property and the technicians of “Barone Ricasoli S.p.A. - Società Agricola” for their support in the experimental activity conducted within the winery vineyards.
Al-Ghussain, L. (2019). Global warming: review on driving forces and mitigation. Environmental Progress and Sustainable Energy, 38(1), 13–21). John Wiley and Sons Inc. https://doi.org/10.1002/ep.13041
Arrizabalaga, M., Morales, F., Oyarzun, M., Delrot, S., Gomès, E., Irigoyen, J. J., Hilbert, G. & Pascual, I. (2018). Tempranillo clones differ in the response of berry sugar and anthocyanin accumulation to elevated temperature. Plant Science, 267, 74–83. https://doi.org/10.1016/j.plantsci.2017.11.009
Aydinalp, C. & Cresser, M. S. (2008). The effects of global climate change on agriculture. American-Eurasian Journal of Agricultural & Environmental Sciences, 3(5), 672–676.
Bergqvist, J., Dokoozlian, N. & Ebisuda, N. (2001). Sunlight exposure and temperature effects on berry growth and composition of Cabernet Sauvignon and Grenache in the Central San Joaquin Valley of California. American Journal of Enology and Viticulture, 52(1), 1–7. https://doi.org/10.5344/ajev.2001.52.1.1
Cameron, W., Petrie, P. R. & Barlow, E. W. R. (2022). The effect of temperature on grapevine phenological intervals: Sensitivity of budburst to flowering. Agricultural and Forest Meteorology, 315, 108841. https://doi.org/10.1016/j.agrformet.2022.108841
Castellarin, S. D., Matthews, M. A., Di Gaspero, G. & Gambetta, G. A. (2007). Water deficits accelerate ripening and induce changes in gene expression regulating flavonoid biosynthesis in grape berries. Planta, 227, 101–112. https://doi.org/10.1007/s00425-007-0598-8
Castellarin, S. D., Pfeiffer, A., Sivilotti, P., Degan, M., Peterlunger, E. & Di Gaspero, G. (2007). Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant, Cell & Environment, 30(11), 1381–1399. https://doi.org/10.1111/j.1365-3040.2007.01716.x
Cataldo, E., Eichmeier, A. & Mattii, G. B. (2023). Effects of Global Warming on Grapevine Berries Phenolic Compounds—A Review. Agronomy, 13(9), 2192.
IPCC, 2014: Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, R.K. Pachauri and L.A. Meyer (eds.)]. IPCC, Geneva, Switzerland, 151 pp.
Costantini E.A.C. (ed), 2013. Oltre la zonazione. Beyond zoning. Tre anni di studio al Castello di Brolio / A three years study at Castello di Brolio. Firenze: Edizioni Polistampa.
Dai, Z. W., Vivin, P., Robert, T., Milin, S., Li, S. H. & Génard, M. (2009). Model-based analysis of sugar accumulation in response to source–sink ratio and water supply in grape (Vitis vinifera) berries. Functional Plant Biology, 36(6), 527–540. https://doi.org/10.1071/FP08284
Dalla Marta, A., Grifoni, D., Mancini, M., Storchi, P., Zipoli, G. & Orlandini, S. (2010). Analysis of the relationships between climate variability and grapevine phenology in the Nobile di Montepulciano wine production area. Journal of Agricultural Science, 148(6), 657–666. https://doi.org/10.1017/S0021859610000432
de Cortazar Atauri, I. G., Duchêne, E., Destrac, A., Barbeau, G., De Rességuier, L., Lacombe, T., Parker, A. K., Saurin, N. & Van Leeuwen, C. (2017). Grapevine phenology in France: from past observations to future evolutions in the context of climate change. Oeno One, 51(2), 115–126. https://doi.org/10.20870/oeno-one.2017.51.2.1622
De Orduna, R. M. (2010). Climate change associated effects on grape and wine quality and production. Food Research International, 43(7), 1844–1855.
De Rességuier, L., Pieri, P., Mary, S., Pons, R., Petitjean, T. & Van Leeuwen, C. (2023). Characterisation of the vertical temperature gradient in the canopy reveals increased trunk height to be a potential adaptation to climate change. Oeno One, 57(1), 41–53. https://doi.org/10.20870/oeno-one.2023.57.1.5365
Drappier, J., Thibon, C., Rabot, A. & Geny-Denis, L. (2019). Relationship between wine composition and temperature: Impact on Bordeaux wine typicity in the context of global warming. Critical Reviews in Food Science and Nutrition, 59(1), 14–30. https://doi.org/10.1080/10408398.2017.1355776
Duchêne, E. (2016). How Can Grapevine Genetics Contribute to the Adaptation to Climate Change? OENO One, 50, 113-124. https://doi.org/10.20870/oeno-one.2016.50.3.98
Duchêne, Eric & Schneider, C. (2005). Grapevine and climatic changes: a glance at the situation in Alsace. Agronomy for Sustainable Development, 25(1), 93–99. https://doi.org/10.1051/agro:2004057
Flexas, J., Escalona, J. M. & Medrano, H. (1998). Down-regulation of photosynthesis by drought under field conditions in grapevine leaves. Functional Plant Biology, 25(8), 893–900. https://doi.org/10.1071/PP98054
Focus, O. I. V. (2017). Distribution of the world’s grapevine varieties. International Organisation of Vine and Wine, 54.
Gambetta, G. A., Herrera, J. C., Dayer, S., Feng, Q., Hochberg, U. & Castellarin, S. D. (2020). The physiology of drought stress in grapevine: towards an integrative definition of drought tolerance. Journal of Experimental Botany, 71(16), 4658–4676. https://doi.org/10.1093/jxb/eraa245
Gambetta, G. A. & Kurtural, S. K. (2021). Global warming and wine quality: are we close to the tipping point? Oeno One, 55(3), 353–361. https://doi.org/10.20870/oeno-one.2021.55.3.4774
Hernández-Montes, E., Zhang, Y., Chang, B.-M., Shcherbatyuk, N. & Keller, M. (2021). Soft, sweet, and colorful: Stratified sampling reveals sequence of events at the onset of grape ripening. American Journal of Enology and Viticulture, 72(2), 137–151. https://doi.org/10.5344/ajev.2020.20050
Hewitt, S., Hernández-Montes, E., Dhingra, A. & Keller, M. (2023). Impact of heat stress, water stress, and their combined effects on the metabolism and transcriptome of grape berries. Scientific Reports, 13(1), 9907. https://doi.org/10.1038/s41598-023-36160-x
Intrigliolo, D. S. & Castel, J. R. (2010). Response of grapevine cv.‘Tempranillo’to timing and amount of irrigation: water relations, vine growth, yield and berry and wine composition. Irrigation Science, 28, 113–125. https://doi.org/10.1007/s00271-009-0164-1
Jackson, D. I. & Lombard, P. B. (1993). Environmental and management practices affecting grape composition and wine quality-a review. American Journal of Enology and Viticulture, 44(4), 409–430. https://doi.org/10.5344/ajev.1993.44.4.409
Keller, M. (2020). The science of grapevines. Academic press.
Keller, M., Romero, P., Gohil, H., Smithyman, R. P., Riley, W. R., Casassa, L. F. & Harbertson, J. F. (2016). Deficit irrigation alters grapevine growth, physiology, and fruit microclimate. American Journal of Enology and Viticulture, 67(4), 426–435. https://doi.org/10.5344/ajev.2016.16032
Kliewer, W. M. (1977). Effect of high temperatures during the bloom-set period on fruit-set, ovule fertility, and berry growth of several grape cultivars. American Journal of Enology and Viticulture, 28(4), 215–222. https://doi.org/10.5344/ajev.1977.28.4.215
Kliewer, W. M. & Lider, L. A. (1968). Influence of cluster exposure to the sun on the composition of Thompson Seedless fruit. American Journal of Enology and Viticulture, 19(3), 175–184. https://doi.org/10.5344/ajev.1968.19.3.175
Koch, B. & Oehl, F. (2018). Climate Change Favors Grapevine Production in Temperate Zones. Agricultural Sciences, 9(3), 247–263. https://doi.org/10.4236/as.2018.93019
Medrano, H., Escalona, J. M., Cifre, J., Bota, J. & Flexas, J. (2003). A ten-year study on the physiology of two Spanish grapevine cultivars under field conditions: effects of water availability from leaf photosynthesis to grape yield and quality. Functional Plant Biology, 30(6), 607–619. https://doi.org/10.1071/FP02110
Mori, K., Goto-Yamamoto, N., Kitayama, M. & Hashizume, K. (2007). Loss of anthocyanins in red-wine grape under high temperature. Journal of Experimental Botany, 58(8), 1935–1945. https://doi.org/10.1093/jxb/erm055
Nesto, B. & Di Savino, F. (2016). Chianti Classico: The Search for Tuscany’s Noblest Wine. (Univ of California Press, Ed.).
Ortiz, A. M. D., Outhwaite, C. L., Dalin, C. & Newbold, T. (2021). A review of the interactions between biodiversity, agriculture, climate change, and international trade: research and policy priorities. One Earth, 4(1), 88–101. https://doi.org/10.1016/j.oneear.2020.12.008
Pastore, C., Dal Santo, S., Zenoni, S., Movahed, N., Allegro, G., Valentini, G., Filippetti, I. & Tornielli, G. B. (2017). Whole plant temperature manipulation affects flavonoid metabolism and the transcriptome of grapevine berries. Frontiers in Plant Science, 8, 929. https://doi.org/10.3389/fpls.2017.00929
Peel, M. C., Finlayson, B. L. & Mcmahon, T. A. (2007). Hydrology and Earth System Sciences Updated world map of the Köppen-Geiger climate classification. In Hydrol. Earth Syst. Sci (Vol. 11). www.hydrol-earth-syst-sci.net/11/1633/2007/
Poni, S., Sabbatini, P. & Palliotti, A. (2022). Facing spring frost damage in grapevine: recent developments and the role of delayed winter pruning–a review. American Journal of Enology and Viticulture, 73(4), 211–226. https://doi.org/10.5344/ajev.2022.22011
Reidmiller, D. R., Avery, C. W., Easterling, D. R., Kunkel, K. E., Lewis, K. L. M., Maycock, T. K. & Stewart, B. C. (Eds.). (2018). Impacts, Risks, and Adaptation in the United States: The Fourth National Climate Assessment, Volume II. https://doi.org/10.7930/NCA4.2018
Reynolds, A. G., Pool, R. M. & Matiick, L. R. (1986). Influence of cluster exposure on fruit composition and wine quality of Seyval blanc grapes. Vitis, 25, 85–95. https://doi.org/10.5073/vitis.1986.25.85-95
Ribéreau-Gayon, P., Glories, Y., Maujean, A. & Dubourdieu, D. (2021). Handbook of Enology, volume 2: The chemistry of wine stabilization and treatments. John Wiley & Sons.
Rienth, M., Vigneron, N., Darriet, P., Sweetman, C., Burbidge, C., Bonghi, C., Walker, R. P., Famiani, F. & Castellarin, S. D. (2021). Grape Berry Secondary Metabolites and Their Modulation by Abiotic Factors in a Climate Change Scenario–A Review. In Frontiers in Plant Science (Vol. 12). Frontiers Media S.A. https://doi.org/10.3389/fpls.2021.643258
Rustioni, L., Altomare, A., Shanshiashvili, G., Greco, F., Buccolieri, R., Blanco, I., Cola, G. & Fracassetti, D. (2023). Microclimate of grape bunch and sunburn of white grape berries: effect on wine quality. Foods, 12(3), 621.
Sadras, V. O. & Moran, M. A. (2012). Elevated temperature decouples anthocyanins and sugars in berries of Shiraz and Cabernet Franc. Australian Journal of Grape and Wine Research, 18(2), 115–122. https://doi.org/10.1111/j.1755-0238.2012.00180.x
Scholasch, T. & Rienth, M. (2019). Review of water deficit mediated changes in vine and berry physiology: consequences for the optimization of irrigation strategies. Oeno One. https://doi.org/10.20870/oeno-one.2019.53.3.2407
Spayd, S. E., Tarara, J. M., Mee, D. L. & Ferguson, J. C. (2002). Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. American Journal of Enology and Viticulture, 53(3), 171–182. https://doi.org/10.5344/ajev.2002.53.3.171
Sweetman, C., Deluc, L. G., Cramer, G. R., Ford, C. M. & Soole, K. L. (2009). Regulation of malate metabolism in grape berry and other developing fruits. Phytochemistry, 70(11–12), 1329–1344. https://doi.org/10.1016/j.phytochem.2009.08.006
Tomasi, D., Jones, G. V, Giust, M., Lovat, L. & Gaiotti, F. (2011). Grapevine phenology and climate change: relationships and trends in the Veneto region of Italy for 1964–2009. American Journal of Enology and Viticulture, 62(3), 329–339. https://doi.org/10.5344/ajev.2011.10108
O.I.V. (2020). 2020 Wine production—OIV first estimates. International Organisation of Vine and Wine Paris.
Van Leeuwen, C., Barbe, J. C., Darriet, P., Destrac-Irvine, A., Gowdy, M., Lytra, G., Marchal, A., Marchand, S., Plantevin, M., Poitou, X., Pons, A. & Thibon, C. (2022). Aromatic maturity is a cornerstone of terroir expression in red wine. Oeno One, 56(2), 335–351. https://doi.org/10.20870/oeno-one.2022.56.2.5441
Van Leeuwen, C. & Darriet, P. (2016). The Impact of Climate Change on Viticulture and Wine Quality. Journal of Wine Economics, 11(1), 150–167. https://doi.org/10.1017/jwe.2015.21
Van Leeuwen, C., Destrac-Irvine, A., Dubernet, M., Duchêne, E., Gowdy, M., Marguerit, E., Pieri, P., Parker, A., De Resseguier, L. & Ollat, N. (2019). An update on the impact of climate change in viticulture and potential adaptations. Agronomy, 9(9), 514. https://doi.org/10.3390/agronomy9090514
Van Leeuwen, C., Sgubin, G., Bois, B., Ollat, N., Swingedouw, D., Zito, S. & Gambetta, G. A. (2024). Climate change impacts and adaptations of wine production. Nature Reviews Earth & Environment, 5(4), 258–275. https://doi.org/10.1038/s43017-024-00521-5
Van Leeuwen, C., Trégoat, O., Choné, X., Bois, B., Pernet, D. & Gaudillère, J.-P. (2009). Vine water status is a key factor in grape ripening and vintage quality for red Bordeaux wine. How can it be assessed for vineyard management purposes? Oeno One, 43(3), 121–134. https://doi.org/10.20870/oeno-one.2009.43.3.798
Wickham, H. & Wickham, H. (2016). Data analysis. Springer.
Winkler, A. J. (1974). General viticulture. University of California Press.
Zapata, D., Salazar, M., Chaves, B., Keller, M. & Hoogenboom, G. (2015). Estimation of the base temperature and growth phase duration in terms of thermal time for four grapevine cultivars. International Journal of Biometeorology, 59, 1771–1781. https://doi.org/10.1007/s00484-015-0985-y
Zarrouk, O., Brunetti, C., Egipto, R., Pinheiro, C., Genebra, T., Gori, A., Lopes, C. M., Tattini, M. & Chaves, M. M. (2016). Grape ripening is regulated by deficit irrigation/elevated temperatures according to cluster position in the canopy. Frontiers in Plant Science, 7, 1640. https://doi.org/10.3389/fpls.2016.01640