Nocturnal transpiration of tomato under deficit irrigation in greenhouse conditions
1 Department of Agronomy, Food, Natural Resources, Animals and Environment – DAFNAE, University of Padua, Agripolis Campus, Viale dell’Università 16, Legnaro, PD, Italy
2 Department of Agriculture, Food and Environment (Di3A) – University of Catania, Via S. Sofia 100, 95123, Catania, Italy
*Corresponding author. E-mail: giorgia.raimondi@unipd.it
Abstract. Nocturnal transpiration (E) can reduce water productivity by causing water loss during a period without photosynthetic activity. This study quantifies tomato nocturnal E under greenhouse conditions, comparing two irrigation managements (full irrigation – FI vs. Deficit irrigation – DI) and four fertilization treatments (raw compost vs. sieved compost vs. mineral vs. no fertilization), addressing a key gap in understanding potential inefficiencies in crop water use. Physiological and environmental parameters were monitored weekly from transplanting to harvesting at four different hours each day (06:00, 12:00, 18:00, and 24:00). At harvest, fruit yield, quality, and water productivity were assessed. Only irrigation significantly affected E, with FI plants exhibiting higher daytime E rates (+11–16%) than DI. Stomatal conductance varied by time but was not influenced by irrigation. Nocturnal E persisted at ~12–13% of daytime rates, indicating residual stomatal opening. Under FI, E positively correlated with leaf temperature and vapor pressure deficit, while under DI, E was more influenced by environmental temperature, reflecting tighter environmental control under water stress. Neither irrigation nor fertilization significantly affected total (on average 64.1 Mg ha⁻¹) or marketable fruit yield (about 77.5% of total yield). Water productivity was significantly higher under DI (+14.7%) than FI (21.5 kg m⁻³). DI also increased fruit dry matter content (+6.5%) and slightly lowered fruit pH without affecting total soluble solids, titratable acidity, or electrical conductivity.
Keywords: transpiration, stomatal conductance, water productivity, fruits’ quality, fruits’ yield.
– Nighttime transpiration was ~12–13% of daytime, unaffected by irrigation management
– Deficit irrigation improved water productivity by 14.7% without yield loss
– Deficit irrigation increased fruits dry matter without altering other quality traits
Water is a critical and increasingly limited resource for agricultural production, directly affecting both crop growth and yield. However, the increase of water productivity (WP) remains a major challenge, particularly in the face of climate change, growing food demand, and the increasing frequency of drought events (FAO, 2021; IPCC, 2022; Borin, 2023). Among the physiological processes involved in plant water use, transpiration (E) plays a fundamental role in leaf temperature (Tleaf) regulation and gas exchange, ensuring the uptake of CO₂ necessary for photosynthesis (Taiz and Zeiger, 2015). Stomata regulate the exchange of gases between the leaf’s internal air spaces and the atmosphere, playing a crucial role in balancing CO₂ uptake for photosynthesis with the prevention of excessive water loss. Due to this dual function, they have become a key target in strategies aiming to enhance WP in crops (Nguyen et al., 2023). However, under suboptimal environmental conditions, stomatal behavior can result in unproductive water loss. For instance, stomata may remain partially open during periods of water stress to support photosynthesis, leading to a significant reduction in WP (Flexas et al., 2013). While traditional strategies have focused on steady-state stomatal conductance (gsw), recent attention to stomatal kinetics and responsiveness offers promising alternatives to enhance WP without compromising carbon assimilation (Nguyen et al., 2023).
Efficient irrigation systems aim to align water supply with plant water demand to maximize productivity. Irrigation is commonly based on reference evapotranspiration (ET0) – which depends on environmental variables such as solar radiation, air temperature, vapor pressure deficit (VPD), and crop coefficient (Kc) (Pereira et al., 2025). However, this simple approach can fall short of capturing crop-specific physiological responses and developmental stages, needing adjustment, for example, considering the deficit irrigation (DI) management (Gong et al., 2020). To overcome this limitation, crop E models that incorporate factors such as leaf area index (LAI), stomatal resistance, and crop development stage have been proposed (Choi and Shin, 2020). Quantifying crop evapotranspiration (ETc) has thus become essential for implementing more targeted and water-efficient irrigation strategies (Sharma and Bhambota, 2022).
Since E is closely linked to plant physiology, it can serve as a reliable indicator of crop growth and development. For this reason, designing irrigation strategies based on E models has become an increasingly important approach to improve irrigation efficiency (Jo et al., 2021). Various methodologies have been developed to estimate E, including the Penman–Monteith, Stanghellini, and Priestley–Taylor models, or to measure E through experimental approaches using soil water balance (Strati et al., 2018), gsw via porometers (Toro et al., 2019), sap flow (Lascano et al., 2016), and weighing lysimeters (Choi and Shin, 2020).
Despite these advances, comparatively little attention has been given to the substantial water losses that occur during the night. Recent evidence suggests that nocturnal E, though not associated with carbon assimilation, may account for a considerable fraction of daily water loss. Across a wide range of C₃ and C₄ plant species, nighttime E has been reported to range from 5% to 15% of daytime E rates, with values reaching as high as 30% under specific environmental conditions (Caird et al., 2007a; Fricke, 2019).
Carbon exchange and water vapor loss through E represent the two major mass flow processes in plants during the day. Interestingly, both continue at night to some extent through partially open stomata (Fricke, 2019). Nighttime E has also been observed in crops grown under artificial lighting and in arid field conditions, suggesting that nighttime E is both widespread and environmentally persistent (Resco de Dios et al., 2016; Fricke, 2019).
The physiological role of nocturnal E remains debated. However, Fricke (2019) suggests that it may offer several benefits, including the maintenance of hydraulic conductivity, facilitation of nutrients transport, and preservation of leaf water potential. Nighttime water loss may also facilitate respiratory CO₂ release through open stomata, a process essential for leaf expansion, particularly under stress conditions such as drought and salinity (Fricke, 2019). Under these circumstances, leaf expansion at night may represent a more efficient use of absorbed water compared to daytime, contributing to stress acclimation mechanisms. In Solanum lycopersicum (tomato), for instance, Lanoue et al. (2017) observed a modest but measurable increase in nighttime E (from 22:00 to 06:00), despite the absence of nocturnal lighting. Although this effect was less pronounced than in Eustoma grandiflorum (lisianthus), the results suggest that tomato exhibits circadian regulation of gsw. Notably, despite similar photosynthetic rates, tomato plants acclimated to red-white and red-blue LED lighting showed a reduction in overall water use efficiency by 25% and 31%, respectively, compared to those grown under high-pressure sodium (HPS) lamps. These findings imply that nocturnal E can substantially impact WP, especially under artificial lighting conditions. From an agronomic perspective, nocturnal water loss can lower WP by consuming irrigation resources without contributing to biomass accumulation and carbon assimilation. Nevertheless, variation in nighttime E among species and genotypes suggests opportunities for genetic selection. A survey of wild and cultivated tomato species showed a range of nighttime E from 8% to 33% of daytime values, highlighting substantial intra- and interspecific variability and breeding potential (Caird et al., 2007b).
Despite the economic importance of tomato, quantitative assessments of nocturnal E in this species remain limited, and no threshold values have been proposed for breeding purposes. In particular, the occurrence of nocturnal E that do not contribute to biomass formation may represent a hidden inefficiency in the plant’s water balance.
This study addresses this gap by quantifying nighttime water loss in greenhouse-grown tomato under different fertilization and irrigation management, also assessing the quanti-qualitative tomato response.
2.1. Experimental site and materials
The study was conducted in a tunnel greenhouse at the “L. Toniolo” experimental farm of the University of Padova (45°21’00” N, 11°57’02” E; 7 m a.s.l.) from June to September 2022. The climate of the area is classified as sub-humid, with an average annual temperature of 13.5 °C. The average annual precipitation (1994–2021) is 830 mm, but evapotranspiration typically exceeds precipitation from April to September by approximately 260 mm (Berti et al., 2014). The soil is classified as Fluvi-Calcaric Cambisol (CMcf) with a silty loam texture (IUSS Working Group WRB, 2014). It has a field capacity and wilting point of 34% (v/v) and 13.5% (v/v), respectively, a bulk density of 1.45 Mg m⁻³, and a slightly alkaline pH (approximately 8).
Before transplanting, two soil tillage operations were performed using a rotary tiller. Fertilization was applied between the two tillage operations, followed by the installation of a drip irrigation system and the transplanting of seedlings. One polyethylene drip line was installed for each tomato row. The drip lines (16 mm diameter) had in-line drippers inserted along the pipe at 0.5 m spacing, with a discharge of 1.1 L h-1. Tomato (HEINZ 1281 F1 - Furia Seed) transplanting took place on June 14, 2022, with a planting density of 2.5 plants m⁻², whereas harvesting on September 27, 2022.
The irrigation volume was determined using soil moisture sensors (Teros 10 - METER Group, Inc., Pullman, WA, USA) placed at three different depths (20 cm, 40 cm, and 60 cm). Irrigation was applied when in the soil layer explored by the plants’ roots, remained the 60% of the available water, restoring soil moisture to field capacity. The seasonal irrigation volume was measured using a water meter.
The experiment followed a split-plot design, with two irrigation management as the main factor (plots of 45 m × 4 m each) and five levels of fertilization as the second factor (subplots of 7.5 m × 4 m each), for a total of 10 plots. The main factor included irrigation at 100% ETc (FI) (for a total irrigation volume of 320 mm) and at 75% ETc (DI) (for a total irrigation volume of 240 mm). The fertilization factor included mineral fertilization, raw compost, fractionated compost (<2 mm), and a control without fertilization. The amount of fertilizers applied provided 150 kg N ha⁻¹, 100 kg P₂O₅ ha⁻¹, and 200 kg K₂O ha⁻¹ to the crop. The characteristics of the compost used in the experiment are reported in Table 1.
| Element | Content |
|---|---|
| Total N | 2.0 % |
| Total C | 22.4% |
| P | 6373 mg kg-1 |
| K | 26549 mg kg-1 |
| Cd | 0.74 mg kg-1 |
| Cr | 36.96 mg kg-1 |
| Cu | 104.64 mg kg-1 |
| Pb | 37.37 mg kg-1 |
| Zn | 247.55 mg kg-1 |
From June 28 to September 6, 2022, physiological (E, gsw, quantum yield of photosystem II - PhiPS2, VPDleaf, Tleaf) and environmental (Tref) parameters were manually measured weekly in six plants per plot using a porometer-fluorometer (LI-600, LI-COR Biosciences, Lincoln, NE, USA). Measurements were taken at four different hours of the day (06:00, 12:00, 18:00, and 00:00). The choice to measure six plants per plot was made to allow the assessment of 60 plants (six plants across ten plots) within approximately one hour, thereby minimizing the potential influence of time on the measured parameters. Additionally, leaves’ chlorophyll content (SPAD index) was measured at 12:00 using a portable chlorophyll-meter (SPAD-502, Minolta, Japan).
On September 13, 2022, three plants per plot were sampled to determine the fruit quality by measuring dry matter content after oven drying at 65 °C, total soluble solids (TSS) content (°Brix), electrical conductivity, and pH. Yield was quantitatively assessed two weeks later, on September 27, 2022, by harvesting five plants per plot. At the end of the growing season the WP was calculated using the following equation:
Descriptive statistics were calculated for all datasets to assess the main characteristics of the data distribution. The normality and homoscedasticity of residuals were evaluated using the Shapiro-Wilk test (Shapiro & Wilk, 1965) and the Bartlett test (Snedecor & Cochran, 1989). When these assumptions were not satisfied, Z-score normalization was applied (Cheadle et al., 2003).
A linear model was fitted using the ‘lm()’ function in R software (Bates et al., 2015) to evaluate E as a function of irrigation regime and sampling hour, including their interaction, across the full dataset. The same approach was used to assess crop yield, WP, and fruits’ quality-related parameters – dry matter content, TSS content, electrical conductivity, and pH – as functions of irrigation and sampling hour, including their interaction, using the complete dataset.
Post hoc analyses were conducted using the ‘emmeans’ package (Lenth et al., 2021) to estimate marginal means, in combination with the ‘rstatix’ or ‘multcomp’ packages (Kassambara, 2019), applying the Sidak method for multiple comparisons.
The correlation between all variables analyzed and T was examined by calculating Spearman’s rank correlation coefficients using the R function ‘cor’ with method = “Spearman”.
3.1. Crop’s physiological traits
The analysis of fertilization and irrigation effects revealed that neither fertilization nor the interaction between the two factors significantly influenced E and gsw. Therefore, based on these results, we focused exclusively on irrigation, excluding fertilization from subsequent analyses.
Considering the entire growing cycle, we observed that E varied significantly over time and between irrigation regimes. The analysis of the irrigation management × sampling hour interaction across the full dataset revealed significant effects on E (Figure 1). At 12:00, the highest E rates were recorded, with FI showing significantly higher values (+11.1%) than the DI. At 12:00 the solar radiation usually peaks as well as the VPD, which might have driven stomatal opening and E, as previously suggested (Grossiord et al., 2020). The higher E observed in FI might have been related to the greater water availability and higher gsw (Chaves et al., 2002).
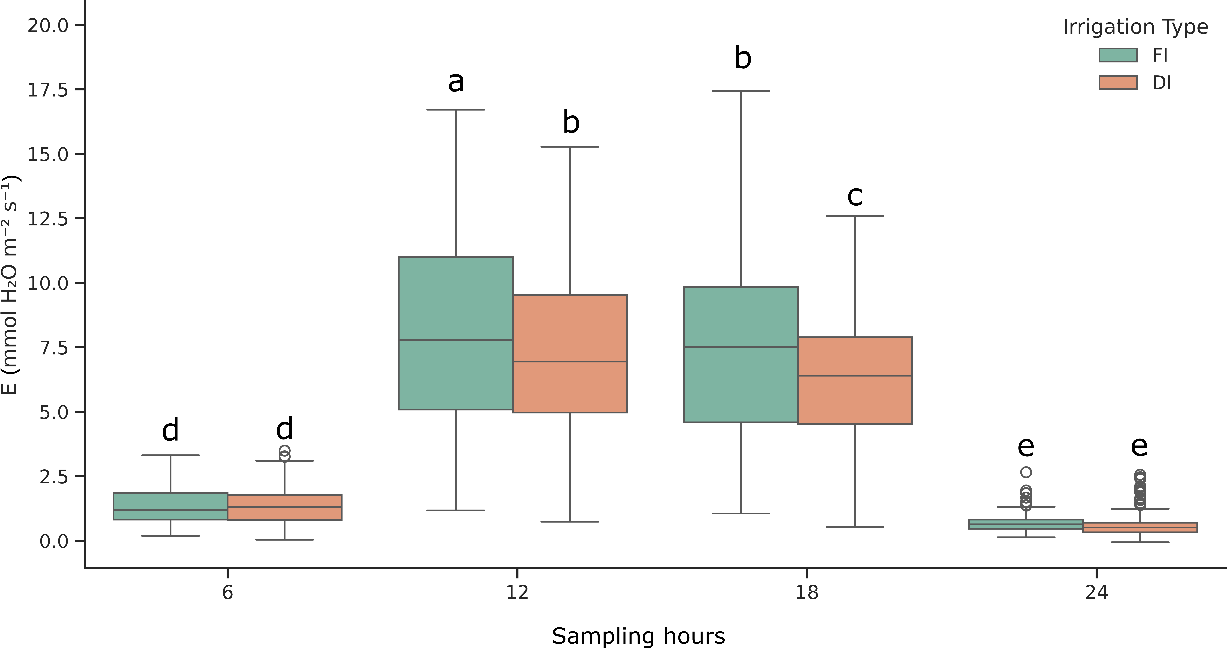
By 18:00, E decreased in both irrigation managements, although it was still significantly higher in FI (+15.8%) than DI. This finding is consistent with the natural decline in light intensity and air temperature. The higher values registered in FI, confirm the relationship between E and water supply even in the afternoon. Throughout the daytime, as reported in literature (Flexas et al., 2006), DI might have induced partial stomatal closure to conserve water, thereby reducing E rate. In addition, the higher relative difference at 18:00 than 12:00 might suggest a cumulative effect of water stress over the day under DI.
No significant effect was instead monitored during the night measurements (6:00 and 24:00) when E was low, but consistently above zero, confirming the findings of previous studies (Caird et al., 2007a; Resco de Dios et al., 2016). The absence of treatment effects at night may suggest that the two compared irrigation managements in our experimental greenhouse conditions have less impact on stomatal behavior during nocturnal periods. On average across the growing season, nighttime measurements showed that E at 06:00 was 1.4 times higher than at 24:00, reaching 0.5 mmol H₂O m⁻² s⁻¹. This might be related to a pre-dawn stomatal opening in anticipation of light, as suggested by the findings of Resco et al. (2009).
On average, the daytime E values (12:00 and 6:00) were 7.7 mmol H2O m-2 s-1 and 6.8 mmol H2O m-2 s-1 for FI and DI managements, respectively. The nighttime E values were the 12.0% and 12.9% of the daytime E values for FI and DI managements, respectively. Our findings agree with Caird et al. (2007b), who measured with a portable photosynthesis system a nocturnal E of 10% of maximal daytime E. The observed nighttime water loss represents a substantial amount of water being lost without simultaneous carbon fixation through photosynthesis. Although this reduction in WP was observed, further physiological assessments are needed to determine possible positive effects, such as lower Tleaf, that may enable faster and more effective recovery from daytime stress, particularly under DI management.
The gsw was significantly influenced by the sampling time, but not by the irrigation management (Table 2). This suggests that the differences observed in E were not only or primarily due to stomatal behavior. It reached its peak at 12:00, with a value of 0.75 mmol H₂O m⁻² s⁻¹. It then decreased by about 25% at 18:00, 75% at 06:00, and 87.5% at 24:00. The higher gsw registered at 12:00, followed by 18:00, confirms the maximal stomatal opening under optimal light conditions, which might explain the corresponding peak in E. The gsw decrease registered at 18:00 coincides with the decrease in E, reinforcing that stomatal aperture is the primary driver of daytime E variation (Flexas et al., 2006). The values of gsw registered at 6:00 and 24:00 reflect a slight residual stomatal conductance corresponding to the non-zero E registered. These findings align with studies showing that nocturnal gsw, while low, can be physiologically meaningful and may contribute to hydraulic redistribution, nutrient uptake, or cooling (Caird et al., 2007b; Resco de Dios et al., 2019).
PhPS2 was significantly affected by the hours, exhibiting the highest values at 24:00 (0.81) and the lowest at 12:00 and 18:00 (0.71). Intermediate values were recorded at 6:00, which did not differ significantly from those at 12:00 and 18:00 (Table 2). No significant differences were found among managements for the leaf SPAD values (56 on average).
| Sampling hours | Physiological parameters | |
|---|---|---|
| gsw | PhiPS2 | |
| 6:00 | 0.20 c | 0.77 ab |
| 12:00 | 0.75 a | 0.71 b |
| 18:00 | 0.58 b | 0.71 b |
| 24:00 | 0.07 d | 0.81 a |
3.2. Physiological parameters’ correlation
The correlation matrices for both night (18:00, 24:00) and day hours (6:00, 12:00) in the two irrigation managements are presented in Figure 2. Only significant correlations (p < 0.05) are described below. E was significantly positively correlated with gsw under both FI and DI managements during both daytime and nighttime. The results are consistent with previous studies showing that increased gsw facilitates water vapor loss from the leaf surface, thereby enhancing plants’ E (Flexas et al., 2012).
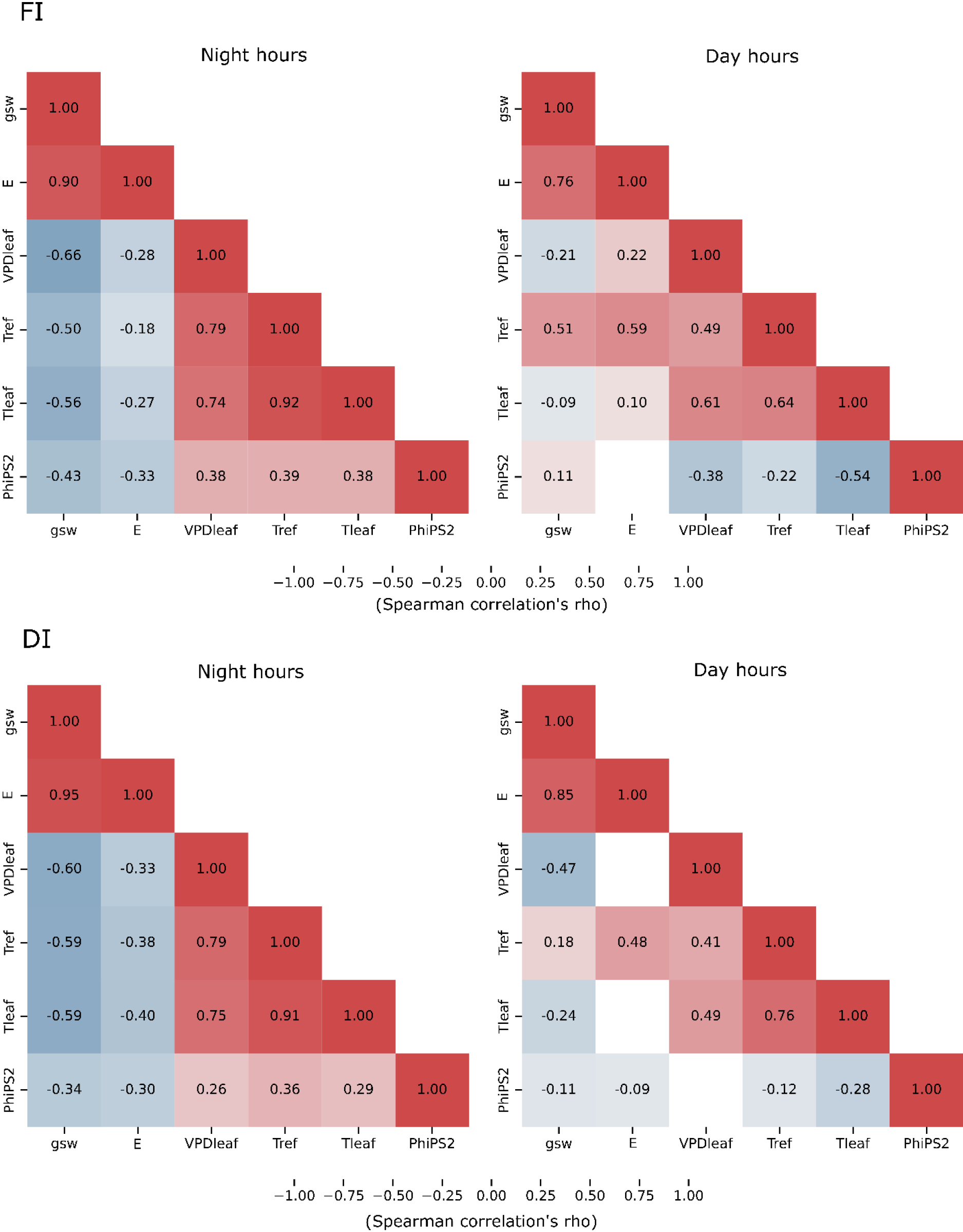
Similarly, a recent study (Savvides et al., 2022) documented a positive relationship between gsw and E under water-stress conditions, although they observed lower E values under DI, a trend not detected in the present study. Although gsw typically decreases at night, the positive nocturnal relationship between gsw and E observed in this study may be explained by residual E, as it did not drop to zero. This pattern aligns with the findings of Caird et al. (2007b). In the nighttime, E was also significantly negatively correlated with VPDleaf under both irrigation managements. This suggests that nocturnal E was more strongly controlled by residual stomatal behavior than by atmospheric water demand. At nighttime, E was significantly negatively correlated with Tleaf and Tref. During daytime, E was positively correlated with Tleaf under FI and with Tref under both irrigation managements. Additionally, E and VPDleaf were significantly negatively correlated at nighttime under both irrigation managements, while a positive correlation was observed under FI during daytime. The correlations between E and Tleaf, Tref, and VPDleaf highlight the role of E in regulating plant thermal balance and vapor pressure dynamics. As reported, E rate drives evaporative cooling, which might affect Tleaf (Gates, 1968). During the night period, without solar radiation input, Tleaf is mainly influenced by Tref, and E-induced evaporative cooling should be minimal. However, in the present study, plants exhibited a non-zero nocturnal E (with related incomplete stomatal closure), which might have caused water loss, consistently with previous observations (Coupel-Ledru et al., 2016), promoting slight evaporative cooling and thereby lowering Tleaf (Caird et al., 2007b). This is consistent with the observed strong positive correlations between Tref and Tleaf during the night under both FI and DI conditions, suggesting that Tref might determine Tleaf at night. At the same time, the cooling effect on Tleaf caused by residual nocturnal E lowers VPDleaf, since VPDleaf depends on air temperature, Tleaf, and ambient humidity, as described by Monteith and Unsworth (2013). Indeed, nocturnal VPDleaf was strongly positively correlated with both Tref and Tleaf, reinforcing the idea that Tleaf plays a key role in nocturnal vapor pressure dynamics in greenhouse conditions (Caird et al., 2007a; Coupel-Ledru et al., 2016). Thus, the nocturnal E, by promoting leaf cooling, might have led to a lower VPDleaf, explaining the observed negative E-VPDleaf correlation under both FI and DI.
During the day, the dynamics shift due to incoming solar radiation, which significantly increases Tleaf. Under FI, high E through open stomata enhances evaporative cooling, helping to counteract heat buildup (Gates, 1968; Jones, 1999). Across the diurnal cycle E and Tleaf are often positively covarying because they share a common driver (increased solar radiation and air temperature): during the midday radiation peak both Tleaf and E can rise together even though E acts to reduce Tleaf relative to the no-transpiration case. This distinction between causal effect (evaporative cooling) and covariation (common forcing by radiation and air temperature) helps explain E–Tleaf positive correlations. Moreover, as daytime temperatures rise and humidity drops, VPDleaf increases, and well-watered plants can respond by increasing stomatal opening, thus sustaining high E. This pattern explains the positive E-VPDleaf correlation observed during daytime under FI. In well-watered, non-stressed conditions, stomata behave passively, responding directly to the evaporative demand driven by increasing VPD rather than actively regulating to conserve water, as described by Monteith (1995) and Jones (2014). In this context, the positive correlations of VPDleaf with Tref and Tleaf observed under FI further support the role of air temperature increases in driving vapor pressure dynamics during the day. Additionally, the strong positive correlation between Tref and Tleaf during the daytime under FI highlights that Tleaf was largely controlled by Tref, even under well-watered conditions.
Regarding Tref, the absence of correlation with E under FI conditions indicates that when water is not limiting, temperature alone is not sufficient to influence E. This suggests that, also under FI, E might be controlled by stomatal and internal plant hydraulic factors by higher extent than external temperatures alone, as reported for water-limited conditions (Chaves et al., 2002; Medrano et al., 2002) where E is primarily regulated by stomatal responses and internal hydraulic constraints (e.g., xylem conductance, water potential), much more than being a direct reaction to external factors like temperature or VPD. Under DI conditions, plants tend to close their stomata to conserve water. This reduces evaporative cooling, making the relationship between E and Tleaf weaker, as registered in the present study where it didn’t result statistically significant. Instead of helping cool the leaf, E might become more influenced by the Tref. This is consistent with the observed positive correlations between Tref and E and the strong positive Tref–Tleaf correlation under daytime DI conditions, suggesting a tighter environmental control of Tleaf and E rates under water deficit. Under DI, plants can be more sensitive to factors like Tref and VPDleaf, which might explain the significant relationships between Tref and E, consistently with the findings of Patakas et al. (2005) reporting that when soil water is limited, plants cannot maintain full stomatal control, making E more tightly linked to external environmental factors like Tref and VPDleaf, especially during daytime when evaporative demand is highest.
E was negatively correlated with PhiPS2 in both irrigation managements during nighttime and in DI during daytime. At night, while the E decreases, in both irrigation managements, PhiPS2 might have undergone basal photochemical and repair activities as previously observed by Flexas et al. (2004) and Baker (2008). Under DI, water stress conditions might have altered photosynthetic efficiency, including the regulation of electron transport rates and photochemical efficiency (Flexas et al., 2004). During daytime, under DI, the negative correlation between E and PhiPS2 might reflect how limited CO₂ uptake under water-limited conditions, together with the light energy, might have caused photochemical impairments, including reduced PhiPS2, as demonstrated in previous studies (Flexas et al., 2004; Chaves et al., 2009; Lawlor and Tezara, 2009). Differently, under FI, adequate water availability might have maintained photosynthetic efficiency, preventing a significant PhiPS2 relationship.
3.3. Crop yield and water productivity
Neither fertilization and irrigation, nor their interaction had a statistically significant effect on total fruits yield, which showed an overall average of 64.1 Mg ha⁻¹. These results are consistent with those reported by Bekele (2017), who found that a 25% reduction in irrigation volume did not significantly affect tomato yield, while improving water productivity.
In the present study, the proportion of marketable yield remained high and comparable between irrigation managements, with values of 77.6% under FI and 77.4% under DI. This suggests that a moderate reduction in irrigation did not compromise fruit yield (Nigatu et al., 2024). Similar findings were reported by Patanè et al. (2011), who demonstrated that DI strategies, including a 50% reduction in ETc applied during part or all of the growing season, did not significantly reduce the marketable yield. Conversely, Lahoz et al. (2016) observed that DI at 75% of ETc led to a 28.2% reduction in water use but also resulted in a 16.4% decrease in marketable yield.
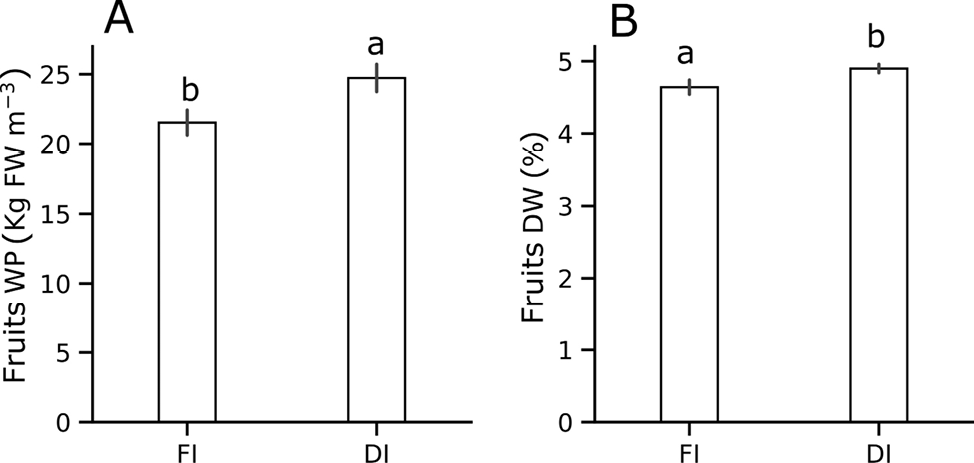
Fruits’ production was significantly enhanced under DI, with a 14.7% increase compared to FI (21.5 kg m⁻³) (Figure 3A). A similar trend was reported by Gragn et al. (2023), who noted a progressive increase in WP as irrigation levels decreased from 100% to 50% ETc. In their study, the highest WP (20.4 kg m⁻³) was achieved at 50% ETc, while the lowest (12.0 kg m⁻³) was recorded at 100% ETc, which was statistically comparable to 75% ETc.
3.4. Marketable fruits’ quality
The application of DI increased the fruits dry matter content (+6.5%) compared to the FI, which recorded 4.9% (Figure 3B). This parameter is particularly relevant for the tomato processing industry, as lower fruit water content is associated with improved processing efficiency (Xu et al., 2024). Instead, no statistically significant differences were observed between irrigation managements for TSS, titratable acidity, or electrical conductivity, with average values of 4.2 °Brix, 6.16, and 4.24 µS/cm, respectively. However, fruits pH was slightly but consistently lower under DI (4.1) than under FI (4.3). Previous studies have highlighted the potential of regulated DI to enhance fruit quality in processing tomato by increasing TSS and other compositional attributes (Xu et al., 2024). For instance, Lahoz et al. (2016) reported an 8.4% increase in TSS and a 2.4% rise in the Hunter a/b ratio, an indicator of improved fruit redness, under DI. However, they did not observe significant changes in pH, contrasting with the slight decrease detected in our study. Our results agree with the findings of Zhang et al. (2017), who found no significant differences in TSS between 70% and 100% ETc irrigation managements, with values ranging from 5.78% to 5.62%. Their findings suggest that moderate water reductions can conserve resources without compromising key fruit quality traits. These improvements are particularly important for the processing sector, as elevated TSS levels contribute to increased product yield and reduced processing costs (Johnstone et al., 2005).
Nocturnal E in tomato plants, although of low magnitude, was consistently detected at 24:00 and 06:00 under both FI and DI, indicating that nocturnal water loss is not negligible. This residual E, likely driven by incomplete stomatal closure, represents a hidden component of the crop’s water balance. Its potential physiological roles, such as contributing to nocturnal leaf cooling, warrant further investigation.
Irrigation management significantly affected daytime E but not nocturnal values. FI resulted in higher E and gsw during peak irradiance (12:00 and 18:00), reflecting passive stomatal behavior under high atmospheric evaporative demand. In contrast, DI induced partial stomatal closure and reduced E, particularly in the afternoon, showing greater dependence on Tref due to limited stomatal control.
DI improved WP by 14.7% without reducing yield. Furthermore, DI enhanced fruit dry matter content by 6.5%, an important quality attribute for processing tomato, without negatively affecting TSS, acidity, or electrical conductivity. These results highlight the potential of moderate water-saving irrigation strategies to improve WP and fruit quality without yield penalties.
This experiment was supported by the project “Deficit irrigation del pomodoro da industria nell’areale veneto” (prot. BIRD 227047) funded by the University of Padova - Department of Agronomy Food Natural resources Animals and Environment (DAFNAE).
Baker, N. R. (2008). Chlorophyll fluorescence: a probe of photosynthesis in vivo. Annu. Rev. Plant Biol., 59(1), 89-113.
Barbour, M. M., & Buckley, T. N. (2007). The stomatal response to evaporative demand persists at night in Ricinus communis plants with high nocturnal conductance. Plant, Cell & Environment, 30(6), 711-721.
Bekele, S. (2017). Response of tomato to deficit irrigation at Ambo, Ethiopia. Journal of Natural Sciences Research, 7(23).
Berti, A., Tardivo, G., Chiaudani, A., Rech, F., & Borin, M. (2014). Assessing reference evapotranspiration by the Hargreaves method in north-eastern Italy. Agricultural Water Management, 140, 20-25.
Borin, M. (2023). A wise irrigation to contribute to integrated water resource management. Italian Journal of Agrometeorology, (2), 5-19.
Caird, M. A., Richards, J. H., & Donovan, L. A. (2007a). Nighttime stomatal conductance and transpiration in C3 and C4 plants. Plant Physiology, 143(1), 4–10. https://doi.org/10.1104/pp.106.092940
Caird, M.A., Richards, J.H., & Hsiao, T.C. (2007b). Significant transpirational water loss occurs throughout the night in field-grown tomato. Functional Plant Biology, 34(3), 172–177.
Chaves, M. M., Pereira, J. S., Maroco, J., Rodrigues, M. L., Ricardo, C. P. P., Osório, M. L., Carvalho, I., Faria, T., Pinheiro, C. (2002). How plants cope with water stress in the field? Photosynthesis and growth. Annals of botany, 89(7), 907.
Chaves, M. M., Flexas, J., & Pinheiro, C. (2009). Photosynthesis under drought and salt stress: regulation mechanisms from whole plant to cell. Annals of botany, 103(4), 551-560.
Cheadle, C., Vawter, M. P., Freed, W. J., & Becker, K. G. (2003). Analysis of microarray data using Z score transformation. The Journal of molecular diagnostics, 5(2), 73-81.
Choi, Y. B., & Shin, J. H. (2020). Development of a transpiration model for precise irrigation control in tomato cultivation. Scientia Horticulturae, 267, 109358.
Coupel-Ledru, A., Lebon, E., Christophe, A., Gallo, A., Gago, P., Pantin, F., ... & Simonneau, T. (2016). Reduced nighttime transpiration is a relevant breeding target for high water-use efficiency in grapevine. Proceedings of the National Academy of Sciences, 113(32), 8963-8968.
FAO (2021). The State of the World’s Land and Water Resources for Food and Agriculture – Systems at breaking point. Food and Agriculture Organization of the United Nations.
Flexas, J., Bota, J., Cifre, J., Mariano Escalona, J., Galmés, J., Gulías, J., Medrano, H. (2004). Understanding down‐regulation of photosynthesis under water stress: future prospects and searching for physiological tools for irrigation management. Annals of applied Biology, 144(3), 273-283.
Flexas J., Bota J., Galmés J., Medrano H. & Ribas-Carbó M. (2006). Keeping a positive carbon balance under adverse conditions: responses of photosynthesis and respiration to water stress. Physiologia Plantarum, 127, 343–352.
Flexas, J., Barbour, M. M., Brendel, O., Cabrera, H. M., Carriquí, M., Díaz-Espejo A., Douthe C., Dreyer E., Juan F. P., Gago J., Gallé A., Galmés J., Kodama N., Medrano H., Niinemets Ü., Peguero-Pina J. J., Pou A., Ribas-Carbó M., Tomás M., Tosens T., Warren, C. R. (2012). Mesophyll diffusion conductance to CO2: an unappreciated central player in photosynthesis. Plant Science, 193, 70-84.
Flexas J., Niinemets Ü., Gallé A., Barbour M.M., Centritto M., Diaz-Espejo A., Medrano H. (2013) Diffusional conductances to CO2 as a target for increasing photosynthesis and photosynthetic water-use efficiency. Photosynthesis Research, 117, 45–59.
Fricke, W. (2019). Night-time transpiration – favouring growth? Trends in Plant Science, 24(4), 311–317.
Gates, David Murray. “Energy exchange in the biosphere.” (1968): 33-43.
Gong, X., Qiu, R., Sun, J., Ge, J., Li, Y., & Wang, S. (2020). Evapotranspiration and crop coefficient of tomato grown in a solar greenhouse under full and deficit irrigation. Agricultural Water Management, 235, 106154.
Gragn, T., Mamo, A., & Wolde, O. (2023). Evaluation of tomato (Solanum Lycopersicum L.) response to deficit irrigation at Adola District, Guji Zone, Southern Ethiopia. Science Research, 11(5), 117-122.
Grossiord, C., Buckley, T. N., Cernusak, L. A., Novick, K. A., Poulter, B., Siegwolf, R. T. W., Sperry, J. S., & McDowell, N. G. (2020). Plant responses to rising vapor pressure deficit. New Phytologist, 226(6), 1550-1566
IPCC (2022). Climate Change 2022: Impacts, Adaptation and Vulnerability. Working Group II Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change.
IUSS Working Group, (2014). World reference base for soil resources 2014. International soil classification system for naming soils and creating legends for soil maps.
Jo, W. J., & Shin, J. H. (2021). Development of a transpiration model for precise tomato (Solanum lycopersicum L.) irrigation control under various environmental conditions in greenhouse. Plant Physiology and Biochemistry, 162, 388-394.
Johnstone, P. R., Hartz, T. K., LeStrange, M., Nunez, J. J., & Miyao, E. M. (2005). Managing fruit soluble solids with late-season deficit irrigation in drip-irrigated processing tomato production. HortScience, 40(6), 1857-1861.
Jones, H. G. (1999). Use of infrared thermometry for estimation of stomatal conductance as a possible aid to irrigation scheduling. Agricultural and forest meteorology, 95(3), 139-149.
Jones, H. G. (2014). Plants and microclimate: a quantitative approach to environmental plant physiology. Cambridge university press.
Lahoz, I., Pérez-de-Castro, A., Valcárcel, M., Macua, J. I., Beltrán, J., Roselló, S., & Cebolla-Cornejo, J. (2016). Effect of water deficit on the agronomical performance and quality of processing tomato. Scientia Horticulturae, 200, 55-65.
Lanoue, J., Leonardos, E. D., Ma, X., & Grodzinski, B. (2017). The Effect of Spectral Quality on Daily Patterns of Gas Exchange, Biomass Gain, and Water-Use-Efficiency in Tomatoes and Lisianthus: An Assessment of Whole Plant Measurements. Frontiers in Plant Science, 8, 1076. https://doi.org/10.3389/fpls.2017.01076
Lascano, R. J., Goebel, T. S., Booker, J., Baker, J. T., & Gitz III, D. C. (2016). The stem heat balance method to measure transpiration: Evaluation of a new sensor. Agricultural Sciences, 7(9), 604-620.
Lawlor, D. W., Tezara, W. (2009). Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: a critical evaluation of mechanisms and integration of processes. Annals of botany, 103(4), 561-579.
Medrano, H., Escalona, J. M., Bota, J., Gulías, J., & Flexas, J. (2002). Regulation of photosynthesis of C3 plants in response to progressive drought: stomatal conductance as a reference parameter. Annals of botany, 89(7), 895-905.
Monteith, J.L. (1995). “A reinterpretation of stomatal responses to humidity.” Plant, Cell & Environment, 18(4), 357–364.
Monteith, J.L., Unsworth, M.H. (2013). Principles of Environmental Physics (4th ed.). Academic Press.
Nguyen, T. B. A., Lefoulon, C., Nguyen, T. H., Blatt, M. R., & Carroll, W. (2023). Engineering stomata for enhanced carbon capture and water-use efficiency. Trends in Plant Science, 28(11), 1290-1309.
Nigatu, B., Getu, D., Getachew, T., & Getaneh, B. (2024). Effect of irrigation regime on yield component and water use efficiency of tomato at Ataye irrigation scheme, Ataye Ethiopia. Italian Journal of Agrometeorology, (2), 89-100.
Patakas, A., Noitsakis, B., & Chouzouri, A. (2005). Optimization of irrigation water use in grapevines using the relationship between transpiration and plant water status. Agriculture, Ecosystems & Environment, 106(2-3), 253-259.
Patanè, C., Tringali, S., & Sortino, O. (2011). Effects of deficit irrigation on biomass, yield, water productivity and fruit quality of processing tomato under semi-arid Mediterranean climate conditions. Scientia Horticulturae, 129(4), 590-596.
Resco, V., Hartwell, J., & Hall, A. (2009). Ecological implications of plants’ ability to tell the time. Ecology letters, 12(6), 583-592.
Resco de Dios, V., Loik, M. E., Smith, R., Aspinwall, M. J., & Tissue, D. T. (2016). Genetic variation in circadian regulation of nocturnal stomatal conductance enhances carbon assimilation and growth. Plant, Cell & Environment, 39(1), 3-11.
Sánchez-Rodríguez, E., Rubio-Wilhelmi, M. M., Cervilla, L. M., Blasco, B., Rios, J. J., Rosales, M. A., Romero, L., & Ruiz, J. M. (2010). Genotypic differences in some physiological parameters symptomatic for oxidative stress under moderate drought in tomato plants. Plant Science, 178(1), 30–40. https://doi.org/10.1016/j.plantsci.2009.10.002
Pereira, L.S., Allen, R.G., Paredes, P., López-Urrea, R., Raes, D., Smith, M., Kilic, A. & Salman, M. 2025. Crop evapotranspiration – Guidelines for computing crop water requirements. Second edition, revised 2025. FAO Irrigation and Drainage Paper, No.56 Rev.1. Rome, FAO. https://doi.org/10.4060/cd6621en
Savvides, A. T., Trouvelot, S., Ruelle, P., & Villemant, C. (2022). Challenging the water stress index concept — A synthesis of plant gas-exchange responses under drought and salinity. Plant, Cell & Environment, 45(8), 2302-2319.
Sharma, V., & Bhambota, S. (2022). Strategies to Improve Crop-Water Productivity. In Food, Energy, and Water Nexus: A Consideration for the 21st Century (pp. 149-172). Cham: Springer International Publishing.
Strati, V., Albéri, M., Anconelli, S., Baldoncini, M., Bittelli, M., Bottardi, C., Mantovani, F. (2018). Modelling soil water content in a tomato field: proximal gamma ray spectroscopy and soil–crop system models. Agriculture, 8(4), 60.
Toro, G., Flexas, J., & Escalona, J. M. (2019). Contrasting leaf porometer and infra-red gas analyser methodologies: an old paradigm about the stomatal conductance measurement. Theoretical and Experimental Plant Physiology, 31(4), 483-492.
Xu, J., Li, X., Wan, W., Zhu, X., Li, C., Zhao, X., Pang, S., Diao, M. (2024). Impact of regulated deficit irrigation on the dynamics of quality changes in processing tomato fruits during ripening. Agricultural Water Management, 304, 109068.
Zhang, H., Xiong, Y., Huang, G., Xu, X., & Huang, Q. (2017). Effects of water stress on processing tomatoes yield, quality and water use efficiency with plastic mulched drip irrigation in sandy soil of the Hetao Irrigation District. Agricultural Water Management, 179, 205-214.